概要
理化学研究所(理研)開拓研究所岩崎RNAシステム生化学研究室の岩崎信太郎主任研究員、脇川大誠リサーチアソシエイト、水戸麻理テクニカルスタッフⅠ、山城はるな特別研究員(研究当時)、戸室幸太郎大学院生リサーチ・アソシエイト、七野悠一上級研究員(研究当時、現筑波大学医学医療系教授)、東京大学大学院理学系研究科の濡木理教授、伊藤弓弦准教授、安藤佑真大学院生、同大学大学院工学系研究科の鈴木勉教授、長尾翌手可講師、東北大学加齢医学研究所の魏范研教授、谷春菜助教、熊本大学大学院生命科学研究部の富澤一仁教授、中條岳志准教授らの共同研究グループは、ミトコンドリア[1]内で行われるタンパク質合成(翻訳[2])の動態(ダイナミクス)を高精度に観測する新しい手法を開発し、ミトコンドリア翻訳[3]の複雑な動態、疾患での制御不全などを明らかにしました。
本研究成果は、ヒトのエネルギー代謝を担うミトコンドリアの仕組みの理解や、ミトコンドリア病[4]などミトコンドリア翻訳異常に関わる疾患の理解につながるものと期待されます。
ヒトの細胞の中には細胞質で起こる翻訳以外にもミトコンドリアの内部で起こる翻訳が存在します。ミトコンドリア内の翻訳を網羅的かつ高解像度に多検体で解析する手法は、望まれつつもこれまで存在しませんでした。
今回、共同研究グループは、新たに「MitoIP-Thor-Ribo-Seq法[5]」という手法を開発し、ミトコンドリア翻訳速度の計測や、ミトコンドリアtRNA(mt-tRNA)[6]修飾による翻訳の促進効果、ミトコンドリア病患者の細胞での翻訳制御不全といった、複雑な翻訳動態を明らかにしました。
本研究は、科学雑誌『Molecular Cell』オンライン版(11月12日付:日本時間11月13日)に掲載されました。

MitoIP-Thor-Ribo-Seq法によるミトコンドリア翻訳のダイナミクスと複雑性の解明
背景
DNAに記された遺伝情報は、メッセンジャーRNA(mRNA)[7]、タンパク質と形を変え、機能を果たします。中でもmRNAからタンパク質がつくられる過程は「翻訳」と呼ばれ、リボソーム[8]によって行われる重要なステップです。この翻訳は、一般的に細胞質という区画で行われますが、これに加えてミトコンドリアと呼ばれる細胞内小器官内で行われる特殊な翻訳(ミトコンドリア翻訳)が存在します。
ミトコンドリアは、細胞内の代謝におけるエネルギーのやりとりを仲立ちするアデノシン三リン酸(ATP)の主要な供給源であり、いわば細胞内のエネルギー工場です。ミトコンドリアは、進化的には細菌に端を発するために独自のゲノムDNA(ミトコンドリアDNA)を持っており、このミトコンドリアDNAにはATP産生に必要な13種のタンパク質の配列情報が記されています。従って、このエネルギー工場内で営まれる翻訳は、エネルギー産生に必須な重要な反応であり、その異常はミトコンドリア病などの疾患につながります。
ミトコンドリア翻訳は、ミトコンドリアDNAから写し取られたmRNAを専用のリボソーム(ミトコンドリアリボソーム)によって読み解くことで行われます。それ故、この翻訳の仕組みは細胞質での翻訳と全く異なります。しかし、細胞質翻訳が大きなバックグラウンドとなる中でミトコンドリア翻訳のみを取り出して解析することは難しく、その詳細な仕組みや全体像はほとんど分かっていませんでした。
近年、mRNA上のリボソームの翻訳動態を網羅的に解析する手法として「Ribo-Seq法[9]」が広く用いられています。Ribo-Seq法では、リボソームフットプリントと呼ばれる短いRNA断片を生成・回収し、次世代シーケンサーでその配列を読み解くことで、mRNA上の「どこに」、「どのくらい」リボソームが存在しているのかといった情報を網羅的に得ることができます(図1A)。この手法では、細胞質翻訳とミトコンドリア翻訳を同時に解析できます(図1A)。しかし、ミトコンドリア翻訳に関する情報は限られるため、ミトコンドリアリボソームの翻訳動態を高解像度に解析することが難しいという課題がありました(図1B)。
そこで共同研究グループは、ミトコンドリア翻訳の解析に特化した新規の手法を開発し、ミトコンドリア翻訳の仕組みの詳細を明らかにしようとしました。

図1 Ribo-Seq法とその課題
(A)Ribo-Seq法の概要。RNase処理:mRNAを分解する酵素で処理すること。オープンリーディングフレーム:mRNAの開始コドン(塩基3個の配列)から終止コドンまでの塩基配列。
(B)Ribo-Seq法によって得られるフットプリントの割合。細胞質リボソームフットプリントに比べ、ミトコンドリアリボソームフットプリントは圧倒的に少ない。
研究手法と成果
共同研究グループは既存のRibo-Seq法を改変して「MitoIP-Thor-Ribo-Seq法」を開発しました。MitoIP-Thor-Ribo-Seq法では、免疫沈降法[10]を用いてミトコンドリアを単離し、ミトコンドリアリボソームを濃縮した状態で、Ribo-Seq法を行います(図2A)。これにより、ミトコンドリアリボソームの翻訳動態を1コドン(塩基3個の配列)分解能で解析することを可能としました(図2B)。
この手法では、ミトコンドリアに局在するタンパク質であるTOM22に対する抗体を用いた免疫沈降(MitoIP)により、ミトコンドリアリボソームを迅速に濃縮します(図2A)。この濃縮を経ることで、回収されるRNAの量は少なくなってしまいますが、RNAを基にRNAを増幅する「Thor[11]」と呼ばれる技術を組み合わせることで、少量サンプルから安定してRibo-Seq法を行うことができます(図2C)。

図2 MitoIP-Thor-Ribo-Seq法の開発
(A)ミトコンドリア免疫沈降法によるミトコンドリアリボソームの濃縮。浸透圧の低い低張液によって細胞膜をやさしく破砕し、細胞内のミトコンドリアを取り出す。その後、TOM22抗体を付着させた磁気ビーズによって、ミトコンドリアを免疫沈降する。
(B)Ribo-Seq法(左)とMitoIP-Ribo-Seq法(右)におけるmRNA上のミトコンドリアリボソームフットプリント数。
(C)Thor-Ribo-Seq法の概要。リンカー:架橋剤、T7:T7 RNAポリメラーゼ、in vitro:試験管内、App:5'-Adenylated phosphate。リンカーライゲーションによって、リボソームフットプリントにT7プロモーターを持ったバーコード配列を付与することで、サンプルの目印化と増幅を可能とする。
共同研究グループは、MitoIP-Thor-Ribo-Seq法、ミトコンドリアリボソームランオフアッセイ法[12]、さらにRibo-Calibration法[13]という手法を組み合わせることで、mRNA上のミトコンドリアリボソーム数や翻訳伸長[14]速度、翻訳開始[15]頻度といった動的なパラメーターを一挙に測定することに成功しました(図3)。これらの手法によって、従来推定不可能だったミトコンドリア翻訳の絶対速度が初めて明らかになりました。その結果、ヒト腎由来HEK293細胞では、翻訳伸長速度が1秒当たり約0.5コドン、翻訳開始は平均435秒に1回程度という極めてゆっくりとしたプロセスであることが判明しました。実際に、これらの翻訳速度はHEK293細胞内の細胞質翻訳と比較すると、伸長速度は約8分の1、開始速度は約20分の1であることが分かりました。また、筋芽細胞であるマウスC2C12細胞でも同様にミトコンドリア翻訳速度を測定したところ、伸長速度が1秒当たり約1.0コドン、翻訳開始は平均173秒に1回とHEK293細胞と比較すると2倍ほど速く、細胞種によってミトコンドリア翻訳動態が大きく異なることも示されました(図3)。
図3 ミトコンドリア翻訳速度の多様性
ヒト腎由来のHEK293細胞(左)とマウスの筋芽細胞C2C12細胞(右)におけるミトコンドリア翻訳速度。
さらに、共同研究グループはmt-tRNAのアンチコドン(リボソームに供給されるアミノ酸の種類に応じた特定の塩基3個の配列)ループに存在する修飾が翻訳に与える影響を網羅的に調べました。mt-tRNAは末端に特定のアミノ酸を結合し、アンチコドンを用いてmRNAのコドンを認識することで、ミトコンドリアリボソームに正しいアミノ酸を供給し、翻訳伸長を可能とします。本研究では、特に読み取りに重要なウォブル位置(34位)に存在する修飾や、アンチコドンの隣の位置(37位)の修飾を欠損(ノックアウト:KO)させた細胞を用いて、MitoIP-Thor-Ribo-Seqを行いました(図4A)。解析の結果、修飾の欠損によって特定のコドンにおいてミトコンドリアリボソームが停滞しやすくなるなど、修飾が翻訳の滑らかさや読み取り精度に大きく関与していることが網羅的に明らかになりました(図4B)。
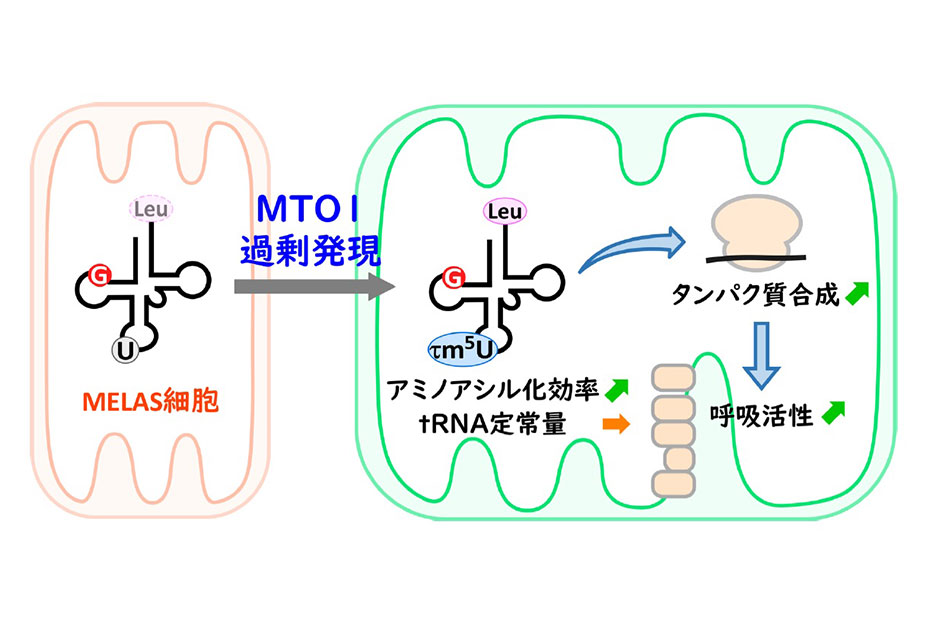
mt-tRNA修飾の異常は、ミトコンドリア病とも密接な関連を持っています。実際に脳卒中様エピソード(MELAS)[16]において最も頻発する原因は、mt-tRNAの病原性点突然変異(A3243G変異)です。この変異によって5-タウリノメチルウリジン(τm5U)(図4A)修飾率が低下し、ミトコンドリア翻訳異常を引き起こすことが知られています。共同研究グループは、MitoIP-Thor-Ribo-Seqを用いて、A3243G変異を持つMELAS患者筋芽細胞でのミトコンドリア翻訳動態を調べました。解析の結果、患者細胞では「U(ウラシル)U(ウラシル)G(グアニン)」コドン上で特異的にミトコンドリアリボソームが停滞していることが明らかになりました(図4C)。さらに、UUGコドンでの顕著なリボソーム停滞は、後続のリボソームの衝突、すなわちリボソーム渋滞を引き起こすことが示されました(図4D)。

図4 mt-tRNA修飾の網羅的解析とMELAS患者細胞でのミトコンドリアリボソーム渋滞
(A)mt-tRNAのウォブル位置(34位)と37位における修飾とその修飾を付与する酵素。ORF:オープンリーディングフレーム。34位の5-ホルミルシチジン(f5C)修飾、5-タウリノメチルウリジン(τm5U)修飾、5-タウリノメチル-2-チオウリジン(τm5s2U)修飾、キューオシン(Q)修飾、37位のN6-スレオニルカルバモイルアデノシン(t6A)修飾の欠損が翻訳伸長に与える影響をMitoIP-Thor-Ribo-Seq法で解析した。
(B)mt-tRNA修飾酵素欠損細胞株における特定コドン上でのミトコンドリアリボソーム停滞の模式図。A、P、EはそれぞれA site、P site、E siteを示す。
(C)健常筋芽細胞およびA3243G変異を持つMELAS患者筋芽細胞におけるUUGコドン(左)、UUA(アデニン)コドン(右)周辺のミトコンドリアリボソーム占有率。
(D)A3243G変異MELAS患者筋芽細胞におけるUUGコドン上でのミトコンドリアリボソーム渋滞の模式図。A、P、EはそれぞれA site、P site、E siteを示す。
今後の期待
MitoIP-Thor-Ribo-Seq法は、ミトコンドリア翻訳の動態を網羅的かつ高解像度に多検体で測定できる新たな技術です。今回明らかになったミトコンドリア翻訳速度の多様性やmt-tRNA修飾の重要性は、ミトコンドリア病や加齢に伴うエネルギー代謝の機能不全や、その細胞種・組織不均一性の理解につながると期待されます。また、本手法はミトコンドリア翻訳異常を発端とするさまざまな疾患の治療標的の探索、ミトコンドリア翻訳の活性化を目的とする創薬研究にも応用が見込まれます。今後、さまざまな細胞や組織に本手法を適用することで、ミトコンドリア翻訳の多様性や調節メカニズムを解き明かし、エネルギー代謝の理解を大きく前進させることが期待されます。
論文情報
<タイトル>
Monitoring the complexity and dynamics of mitochondrial translation
<著者名>
Taisei Wakigawa, Mari Mito, Yushin Ando, Haruna Yamashiro, Kotaro Tomuro, Haruna Tani, Kazuhito Tomizawa, Takeshi Chujo, Asuteka Nagao, Takeo Suzuki, Osamu Nureki, Fan-Yan Wei, Yuichi Shichino, Yuzuru Itoh, Tsutomu Suzuki, Shintaro Iwasaki
<雑誌>
Molecular Cell
<DOI>
10.1016/j.molcel.2025.10.022
補足説明
[1] ミトコンドリア
真核細胞内に存在する小器官で、独自のDNAを持ち、アデノシン三リン酸(ATP)を産生する「エネルギー工場」。
[2] 翻訳
メッセンジャーRNA(mRNA)([7]参照)に記された塩基配列をアミノ酸配列へ変換し、リボソーム([8]参照)でアミノ酸を結合してタンパク質を合成する過程。
[3] ミトコンドリア翻訳
ミトコンドリアDNAから写しとられたmRNAからタンパク質を合成する過程。ミトコンドリア内で、専用のミトコンドリアリボソームによって行われる。
[4] ミトコンドリア病
ミトコンドリア機能異常に基づく多臓器疾患の総称。代表的な病例として、脳卒中様エピソード(MELAS)([16]参照)、ミオクローヌスてんかん(MERRF)があり、ミトコンドリア翻訳異常が病因となることが多い。
[5] MitoIP-Thor-Ribo-Seq法
ミトコンドリア免疫沈降法(MitoIP)とRNA増幅法(Thor([11]参照))を組み合わせることでミトコンドリアリボソームの解析に特化したRibo-Seq法([9]参照)。網羅的かつ高精度にミトコンドリア翻訳を解析できる。
[6] ミトコンドリアtRNA(mt-tRNA)
ミトコンドリアDNAにコードされる22種類の転移RNA(tRNA)の総称。ミトコンドリアリボソームにmRNA情報に基づいたアミノ酸を供給し、翻訳伸長([14]参照)を支える。
[7] メッセンジャーRNA(mRNA)
タンパク質のアミノ酸配列情報(コドン)を持つRNA。リボソームによってコドンが読み取られることでタンパク質が合成される。
[8] リボソーム
リボソームRNA(rRNA)とリボソームタンパク質で構成される巨大複合体。
[9] Ribo-Seq法
翻訳装置であるリボソームを抽出し、リボソームと結合しているmRNA配列を同定することで、どの遺伝子がどの程度の効率で翻訳されているかを解析する手法。リボソームは大きな複合体であり、一定のmRNA領域を覆うように結合するため、これらのリボソームとmRNAの複合体をRNA分解酵素で処理すると、リボソームが保護するmRNA断片だけが分解されずに回収される。
[10] 免疫沈降法
特定のタンパク質やタンパク質複合体を抗体によって沈殿させ、目的の分子や小器官を選択的に回収・濃縮する方法。
[11] Thor
T7 RNAポリメラーゼを用いて、RNAを鋳型にRNAを増幅する手法。Ribo-Seq法において、フットプリントの増幅に用いられる。ThorはT7 high-resolution original RNAの略。
[12] ミトコンドリアリボソームランオフアッセイ法
一定時間を置きmRNA上で翻訳反応(ラン/run)しているミトコンドリアリボソームに、翻訳を終結させ、乖離(かいり)(オフ/off)させる実験手法。ミトコンドリア翻訳開始を阻害することで実験が可能になる。
[13] Ribo-Calibration法
絶対的なリボソーム数の測定を可能としたRibo-Seq法の変法。既知のリボソーム数(モル比)から成るmRNA・リボソーム複合体を細胞に添加し、Ribo-Seq法で得られたデータを補正することで、mRNA上で何個のリボソームが翻訳していたのかについて網羅的な情報が得られる。
[14] 翻訳伸長
mRNA上に結合したリボソームが、コドンを認識し、ペプチドを伸長させる反応を指す。
[15] 翻訳開始
リボソームがmRNA上の開始コドン上に結合するまでの一連の素過程(基本的な過程)を指す。
[16] 脳卒中様エピソード(MELAS)
ミトコンドリア病の一種で、エネルギー代謝異常により脳細胞が機能不全に陥ることで発症する。mt-tRNAの病原性点突然変異を原因とすることが多く、ミトコンドリア翻訳に異常を来す。
共同研究グループ
理化学研究所 開拓研究所 岩崎RNAシステム生化学研究室
主任研究員 岩崎信太郎(イワサキ・シンタロウ)
リサーチアソシエイト 脇川大誠 (ワキガワ・タイセイ)
テクニカルスタッフⅠ 水戸麻理 (ミト・マリ)
特別研究員(研究当時) 山城はるな(ヤマシロ・ハルナ)
大学院生リサーチ・アソシエイト 戸室幸太郎(トムロ・コウタロウ)
上級研究員(研究当時) 七野悠一 (シチノ・ユウイチ)
(現 筑波大学 大学院医学医療系 RNA生化学 教授)
東京大学
大学院理学系研究科 生物化学専攻
教授 濡木 理 (ヌレキ・オサム)
准教授 伊藤弓弦 (イトウ・ユヅル)
大学院生 安藤佑真 (アンドウ・ユウシン)
大学院工学系研究科 化学生命工学専攻
教授 鈴木 勉 (スズキ・ツトム)
講師 長尾翌手可 (ナガオ・アステカ)
東北大学 加齢医学研究所 モドミクス医学分野
教授 魏 范研 (ウエイ・フアンイエン)
助教 谷 春菜 (タニ・ハルナ)
熊本大学 大学院生命科学研究部 総合医薬科学部門
教授 富澤一仁 (トミザワ・カズヒト)
准教授 中條岳志 (チュウジョウ・タケシ)
琉球大学 大学院医学研究科 医化学講座
教授 鈴木健夫 (スズキ・タケオ)
研究支援
本研究は、RIKEN Pioneering Projects「Biology of Intracellular Environments(研究分担者:岩崎信太郎、七野悠一)」、RIKEN TRIP initiative(AGIS注)・細胞応答モデル開発)(参画研究者:岩崎信太郎)、日本学術振興会(JSPS)科学研究費助成事業基盤研究(B)「ミトコンドリア特異的リボソームプロファイリング法によるオルガネラ翻訳網羅解析(研究代表者:岩崎信太郎)」「ミトコンドリアのリボソーム形成過程の構造解析(研究代表者:伊藤弓弦)」、同基盤研究(A)「細胞内構造を支えるヘテロポリマー間相互作用の網羅的解析(研究分担者:岩崎信太郎)」、同学術変革領域研究(B)「新規Disome-Seq法:パラメトリックなリボソーム渋滞の網羅的探索(研究代表者:岩崎信太郎)」、同学術変革領域研究(A)「時間タンパク質学:翻訳速度の大規模並列網羅解析(研究代表者:岩崎信太郎)」「APEX-Ribo-Seq:近傍標識による非典型局所翻訳の網羅解析(研究代表者:七野悠一)」、同基盤研究(C)「細胞内顆粒P-bodyによるmRNAの空間的制御を介した細胞周期調節機構(研究代表者:七野悠一)」「非典型局所翻訳を包括的に解明するAPEX-Ribo-Seq法の確立(研究代表者:七野悠一)」、同若手研究「翻訳開始因子eIF4A1とグルタミン代謝による協調的な発現制御機構(研究代表者:七野悠一)」、同研究活動支援スタート支援「ミトコンドリアリボソーム停滞が引き金となる新規mtUPR経路の解明(研究代表者:脇川大誠)」、同特別研究員奨励費「細胞接着によるミトコンドリア翻訳制御の分子メカニズムの解明(研究代表者:脇川大誠)」「クライオ電子トモグラフィーを用いたミトコンドリアタンパク質輸送機構の解明(研究代表者:安藤佑真)、東京大学大学院新領域創成科学研究科挑戦的新領域博士研究助成制度「細胞接着によるミトコンドリア翻訳活性化機構の解明(研究代表者:脇川大誠)」「ミトコンドリアribosomeの衝突を解消する品質管理機構の解明(研究代表者:脇川大誠)」、科学技術振興機構(JST)戦略的創造研究推進事業ERATO「鈴木RNA修飾生命機能プロジェクト(研究総括:鈴木勉)」、日本医療研究開発機構(AMED)生命科学・創薬研究支援基盤事業(BINDS)「クライオ電子顕微鏡による分子・細胞構造解析の支援と高度化(JP23ama121002)」による助成を受けて行われました。
注)AGIS:Advanced General Intelligence for Science Program(AGIS科学研究基盤モデル開発プログラム)
発表者
<発表者> ※研究内容については発表者にお問い合わせください。
理化学研究所 開拓研究所 岩崎RNAシステム生化学研究室
主任研究員 岩崎信太郎 (イワサキ・シンタロウ)
リサーチアソシエイト 脇川大誠 (ワキガワ・タイセイ)
テクニカルスタッフⅠ 水戸麻理 (ミト・マリ)
特別研究員(研究当時) 山城はるな (ヤマシロ・ハルナ)
大学院生リサーチ・アソシエイト 戸室幸太郎 (トムロ・コウタロウ)
上級研究員(研究当時) 七野悠一 (シチノ・ユウイチ)
(現 筑波大学 医学医療系 教授)

東京大学
大学院理学系研究科
教授 濡木 理 (ヌレキ・オサム)
准教授 伊藤弓弦 (イトウ・ユヅル)
大学院生 安藤佑真 (アンドウ・ユウシン)
大学院工学系研究科
教授 鈴木 勉 (スズキ・ツトム)
講師 長尾翌手可 (ナガオ・アステカ)
東北大学 加齢医学研究所
教授 魏 范研 (ウエイ・フアンイエン)
助教 谷 春菜 (タニ・ハルナ)
熊本大学 大学院生命科学研究部
教授 富澤一仁 (トミザワ・カズヒト)
准教授 中條岳志 (チュウジョウ・タケシ)
<発表者のコメント>
ミトコンドリア翻訳の制御メカニズムは、その解析の難しさからまだまだ分かっていないことだらけです。今回私たちが開発したMitoIP-Thor-Ribo-Seq法が、かすみがかったエネルギー工場内をのぞく一助となり、さらなる発見につながることを願っています。(脇川大誠)
プレスリリース本文:PDFファイル
Molecular Cell:https://doi.org/10.1016/j.molcel.2025.10.022
病気の変異を持ったままでも光明が見えてきた ―tRNA修飾酵素でミトコンドリア病の治療を目指す―

小胞体とミトコンドリアが手をつなぐ仕組み ―発達障害の原因解明に期待―
